Популяционная генетика. Факторы, характеризующие популяцию, влияющие на прохождение мутационных штурмов
На эволюционные процессы в популяциях влияют разные факторы: Численный состав «банка рецессивных генов» популяции и «очередь» рецессивных генов на «внедрение». Формирование «подвала», обеспечивающего сохранность малочисленных рецессивных генов в течение многих поколений. Разные проявления «характера» генов, размер популяции. Факторы, влияющие на формирование и сохранение разнообразия рецессивных генов в популяции.

Прежде чем начать новую статью я хочу уточнить некоторые понятия которые я использовал в предыдущих статьях, видимо, не совсем точно и мог затруднить понимание сути читателем. А именно: 1. Я говорил о «близкородственном скрещивании в большой популяции». Это противоречивое высказывание могло сбить с толку. Я имел ввиду скрещивание 2-х обладателей редких одинаковых рецессивных генов в гетерозиготном состоянии. Такое явление, как неоднократно я говорил, чаще всего имеет место при близкородственном скрещивании. Дабы подчеркнуть единство подобных процессов я оба процесса обозвал «близкородственным скрещиванием», но при этом в первом случае добавлял, дабы подчеркнуть различие, «в большой популяции». 2. Я применил два выражения использовавшие однокоренные слова: «доминантные гены» и «доминирующие гены». Доминирующие – в смысле преобладающие, находящиеся в большинстве. Естественно, что и рецессивные гены могут быть доминирующими.
В одной из предыдущих статей[4] в разделе 7 мы коснулись состава банка рецессивных генов, однако, другие задачи статьи не позволили детально рассмотреть, как меняется «поведение» генов в зависимости от «концентрации» «новых» рецессивных генов в популяции. Сейчас, есть смысл подробнее заняться этим вопросом.
1. Численный состав банка рецессивных генов популяции и «очередь» рецессивных генов на «внедрение».
Начать надо с периода «застоя» в эволюционном процессе, характеризуемого относительно стабильным состоянием рецессивных генов в гетерозиготном и в гомозиготном состояниях. Естественный отбор делает своё дело, но этот процесс идёт медленно. Обладатели лучших с «точки зрения» естественного отбора рецессивных генов в гомозиготном состоянии постепенно увеличивают долю в популяции меняя соотношение «новых» мутированных рецессивных генов к старым немутированным. Зная долю «нового» рецессивного гена в популяции, мы можем рассчитать долю генов, находящихся в гетерозиготном и в гомозиготном состояниях. Теория вероятности события даёт нам возможность подсчитать это соотношение, поскольку оно будет определяться вероятностью событий «встречи» двух разных или двух «новых» генов в потомстве. В случае встречи двух «новых» одинаковых мы получаем пару генов в гомозиготном состоянии и тогда они начинают проявлять свои доселе спящие качества, и мы увидим рецессивный ген в работе. Вероятность «встречи» двух одинаковых рецессивных генов можно представить на модели: Представьте, что мы пытаемся достать из мешка с красными и белыми шариками сразу два красных шарика или с начала один, потом второй, что, согласитесь, тоже самое. Всего в мешке 100 шариков. Вероятность, что мы достанем красный шарик в первый раз равна Рк = К. Где К – доля красных шариков в мешке. Вернём извлечённый из мешка шарик обратно в мешок, что бы всегда в мешке было 100 шариков. Вероятность, что второй шарик будет красным, естественно, такая же. Тогда вероятность 2-х событий подряд – 2-х красных шариков, что равнозначно доставанию пары красных шаров будет равна произведению вероятностей, то есть Рк2 = КхК. Это квадратное уравнение и графиком зависимостей вероятности «встречи» от доли гена в популяции будет ветвь параболы.
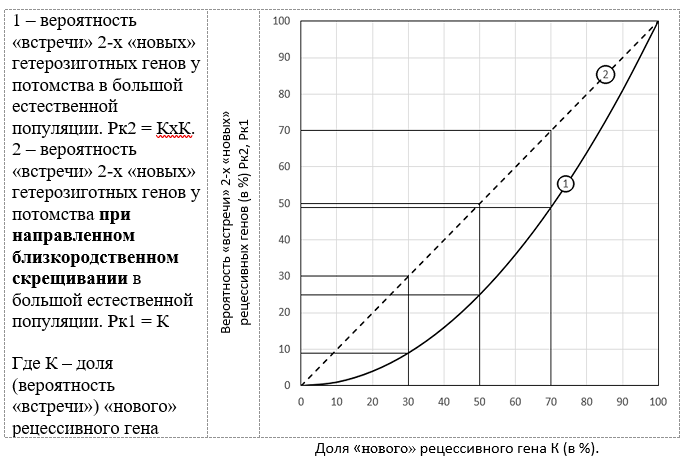
Рисунок 1. Вероятность встречи «новых» рецессивных генов в гетерозиготном состоянии Рк2 у потомства в зависимости от содержания гена К в популяции.
Из графика 1 мы видим, что при низких значениях содержания «нового» рецессивного гена вероятность встречи двух одинаковых генов в гетерозиготном состоянии в большой естественной популяции очень мала: при К = 0,1 (10%), Рк2 = 0,1х0,1 = 0,01 (1%); не трудно посчитать и другие значения вероятности «встречи» 2-х одинаковых генов в зависимости от содержания гена в популяции. Данные сведены в таблицу:
Таблица 1
Вероятность встречи «новых» рецессивных генов в гомозиготном состоянии Рк2 у потомства в зависимости от содержания гена К в популяции
|
Рк1=К, |
0,1 10% |
0,2 20% |
0,3 30% |
0,4 40% |
0,5 50% |
0,6 60% |
0,7 70% |
0,8 80% |
0,9 90% |
1,0 100% |
|
Рк2=КхК, |
0,01 1% |
0,04 4% |
0,09 9% |
0,16 16% |
0,25 25% |
0,36 36% |
0,49 49% |
0,64 64% |
0,81 81% |
1,0 100% |
Я условно разделил этот график на четыре зоны: при доле нового рецессивного гена К от 0 до 0,3 (30%) вероятность «встречи» 2-х одинаковых рецессивных генов меняется от 0 до 0,09 (9%) - это зона «подвала», где встречи маловероятны; при К от 0,3 (30%) до 0,07 (70%), Рк2 меняется от 0,09 (9%) до 0,5 (50%) - это нижняя и верхняя часть средней зоны, где вероятность встречи 2-х «новых» рецессивных генов меньше 0,49 (49%) и, соответственно верхней – при К выше 0,7 (70%) и вероятность образования гомозиготной пары больше 0,5 (50%).
О чём говорят эти цифры? О том, что при низких концентрациях «нового» рецессивного гена в гетерозиготном состоянии при увеличении его концентрации от 0 до 0,1 (10%) вероятность встречи двух одинаковых «новых» генов возрастает от 0 до 0,01 (1%) – незначительно – на 0,01 (1%), в то же время, при увеличении концентрации на ту же долю в 0,1 (10%) при изменении концентрации 0,9 (90%) до 1,0 (100%) вероятность увеличивается на 0,19 (19%). А это значит:
-- «Новые» гены, находящиеся в популяции в небольших количествах, например, до 10% - из них в гомозиготном состоянии будут представлены в количестве до 1%, то есть, большая часть будет представлена в гетерозиготном состоянии и никак не проявят своих рабочих качеств, что делает их недоступными для естественного отбора. И даже при концентрации «нового» рецессивного гена в 30% доля в гомозиготном состоянии составит только 9%. Я, условно, назвал эту зону концентраций «подвалом», где «прячутся» от естественного отбора как отрицательные, так и другие, невостребованные сегодня естественным отбором гены, в том числе, когда-то используемые и потенциально полезные и всякие другие в том числе нейтральные гены. Это «склад» множества мутаций за последние, возможно, несколько миллионов лет. Верхний уровень концентрации определяется количеством мутационных штурмов в которых участвовали эти гены, так как они увеличивают долю редких генов, а также степенью их вредности – вредные уничтожаются естественным отбором. При совсем низких концентрациях вероятность их уничтожения очень мала. Как отрицательные, так и положительные будут храниться в «подвале» и передаваться из поколения в поколение. Они никак не влияют на работающую часть генов - на доминантные и рецессивные в гомозиготном состоянии. Именно разнообразие «подвальных» генов определяет возможность популяции находить спасительные «новые» гены в критических ситуациях. Только при близкородственном скрещивании естественным отбором могут быть «выявлены» «новые» перспективные гены.
-- При распространении «нового» рецессивного гена в популяции в средней части концентраций от 30% до 70% происходит интенсивный процесс естественного отбора. Прежде всего, в эту зону могут попасть со стороны «подвала» отобранные естественным отбором гены после увеличения численности в результате близкородственного скрещивания, с другой - со стороны 100 % доминирования рецессивного гена в гомозиготном состоянии, утратившие своё доминирующее положение в прошлом гены. В этой зоне естественный отбор будет определять динамику изменения концентрации обладателей того или иного гена. Проявляющие положительные качества будут увеличивать численность их обладателей и наоборот. При отсутствии естественного отбора изменений концентраций не будет. Доля их будут сохраняться в последующих поколениях.
-- Следующая зона – от 70% до 100% обладателей «нового» рецессивного гена в популяции. Это зона быстрого достижения доминирования «нового» гена в популяции. И даже при отсутствии естественного отбора направленного на увеличение численности обладателей «нового» гена доля его обладателей в популяции будет увеличиваться.
2. Напомню, что «подвал» банка рецессивных генов составляют гены:
- Положительные, хранящиеся очень давно, но никак не способные по концентрации рецессивного гена в гетерозиготном состоянии войти в среднюю зону (зону эффективного скрещивания в гетерозиготном состоянии)
- Положительные, но утратившие свою актуальность в связи с внедрением более перспективных с точки зрения естественного отбора.
- Разнообразные, в том числе не эффективные и отрицательные, прошедшие мутационные штурмы и набравшие за счёт близкородственного скрещивания минимальное количество – минимальную долю для сохранения в популяции в «подвале».
- Разнообразные недавно мутированные единичные рецессивные гены.
3. Теоретически, «молодые» мутированные гены, как и все другие, представленные малом количеством в гетерозиготном состоянии могут храниться в банке рецессивных генов бесконечно долго в «подвале», хотя, вероятность потери единичных генов больше чем других, но, периодически, проходящие мутационные штурмы увеличивают концентрацию единичных рецессивных генов. Для увеличения концентрации гена не получившего доминирующего распространения в период застоя, ему важно попасть в «бутылочное горлышко», при близкородственном скрещивании, что, конечно же, удаётся не многим, особенно малочисленным «обитателям подвала», а значит, и недавно мутированным – «молодым». У рецессивных генов, получивших большое распространение в популяции в период застоя, большая вероятность попасть в «бутылочное горлышко» при очередном мутационном штурме.
4. Среднюю и верхнюю часть средней зоны графика (рис. 1, кривая 1) определяют результаты прошедшего мутационного штурма и естественного отбора в большой популяции. Ведь мутационный штурм даёт всем рецессивным генам необоснованное «их способностями» увеличение численности и только естественный отбор может одних обладателей рецессивных генов уничтожить, а другим дать размножиться. Именно гены, определяющие способность увеличивать численность популяции в дальнейшем займут среднюю и верхнюю зоны графика. Именно они в период «застоя» смогут завоевать первенство в гомозиготном состоянии.
5. При рассмотрении графика (рис. 1, кривая 1) точку в 50% - (0,5) вероятности встречи рецессивных генов можно рассматривать как переломную – до концентрации «нового» рецессивного гена в 70% - (0,7) больше встреч «нового» гена будет с чужеродным геном, а значит будет больше рождаться детей с гетерозиготным геном в спящем состоянии и при концентрации свыше 70% будет больше рождаться детей с «новым» геном в гомозиготном состоянии, и они будут демонстрировать качество работоспособного гена. Перед вами наглядно представляется «очередь» генов имеющих разную вероятность для «внедрения».
6. Возникает естественный вопрос: проявляют ли свои качества рецессивные гены у отдельных представителей популяции, не набравшие значительной доли в популяции? Конечно же - да. Именно эти гены и отличают нас внутри популяции. Разные проявление отдельных качеств, есть, в том числе, проявление разных рецессивных генов в гомозиготном состоянии. Доминантные гены в меньшей степени обладают разнообразием. Будь они одинаковыми мы бы не видели между нами разницы. В зависимости от доли рецессивного гена в популяции мы можем наблюдать его проявление с той или иной частотой. В расчётах, с вероятностью, нашедшей отражение в графике 1 проявление тех или иных качеств – рецессивного гена в гомозиготном состоянии (см. рис.1).
Обитатели «подвала» имеют наименьшую вероятность «встретиться» и проявить себя. Иногда эти качества касаются видимых изменений наследственности. И эти качества, хранящиеся в виде редких рецессивных генов в гетерозиготном состоянии не проявляемые ни у родителей, ни у известных предков, и вдруг, проявляются у детей в виде неожиданного качества (в гомозиготном состоянии). Этих проявлений не помнят родители у ближайших предков, поскольку они могли быть у очень давних предков, а тут на тебе... они «вылезают». С большей вероятностью можно встретить такие случаи при близкородственном скрещивании. Это явление, как мы видим, носит наследственный характер, однако, поскольку у известных родителям предков «новое» качество не наблюдалось, создаётся впечатление, что это абсолютно новое явление – какой-то не менделевский процесс - «дрейф генов» со стороны. Хотя, как мы видим, всё объяснимо и очень просто. Вот такое проявление исключительной наследственности от далёких предков. Рождались даже псевдо научные теории типа телегонии.
Исключительной наследственностью от далёких предков можно объяснить и проявление у потомства склонности к проявлению нетрадиционной половой ориентации. Правда, никак не могут найти эти гены, отвечающие за нарушение традиционной половой ориентации, хотя, всё чаще однозначно связывают проявление данных качеств с наследственностью.
7. В связи с естественной актуальностью для жизни человека наилучшим образом изучены вопросы «встречи» двух рецессивных генов, вызывающих наследственные заболевания, хотя и любые другие гены, представленные малым количеством ведут себя аналогично. В свете рассматриваемой выше квадратичной зависимости (рис. 1 кривая 1 и таблица 1) вероятности встречи 2-х генов будут определять проявления наследственных заболеваний. Если вероятность встречи 2-х рецессивных генов определяется как Рк2 = КхК. Но, в природе, в определённых условиях, прежде всего, из-за совместного проживания родственников и в человеческих популяциях, когда вступление в брак определяется традициями и другими мотивами (сохранение наследства) возможно скрещивание близких родственников. В таком случае, когда вступающий в брак является носителем рецессивного гена аутосомного заболевания равна вероятность Рк1 = К (см. рис 1, прямая 2). В случае близкородственного скрещивания, для получения гена в гомозиготном состоянии нам уже не нужен второй обладатель. В случае близкородственных скрещиваний потомков мы непременно его получим в следующих поколениях. О чём мы говорили ранее. От скрещивания двух обладателей рецессивных генов в гетерозиготном состоянии мы получим обладателя рецессивного гена в гомозиготном состоянии. Таким образом вероятность встречи в первом приближении будет определяться вероятностью попадания одного красного шара. Графиком Рк1 = К будет прямая 2 соединяющая точки Рк2 = Рк1 = 0 и Рк2 = Рк1 = 100%. Поскольку, вероятность или фактическое проявление наследственного заболевания принято выражать в единицах на сколько то тысяч, например, 1 заболевший на 10000 родившихся детей или 1 на 1000, то в таблице 2 представлена вероятность или частота проявлений заболевания в соответствующих единицах.
Таблица 2
Вероятность или фактическое проявление наследственного заболевания аутосомно рецессивного характера в большой естественной популяции Рк2 и при направленном близкородственном скрещивании Рк1, а также отношение Рк1 / Рк2.
|
Рк2 =КхК, |
0,000001 1 из 1000000 |
0,000005 1 из 200000 |
0,00001 1 из 100000 |
0,00005 1 из 20000 |
0,0001 1 из 10000 |
0,0005 1 из 2000 |
0,001 1 из 1000 |
0,005 1 из 200 |
|
Рк1= К, |
0,001 1 из 1000 |
0,00224 1 из 448 |
0,0032 1 из 316 |
0,0071 1 из 141 |
0,01 1 из 100 |
0,0224 1 из 45 |
0,032 1 из 32 |
0,071 1 из 14 |
|
Рк1/ Рк2=К |
1000 |
448 |
316 |
141 |
100 |
44,8 |
32 |
14,1 |
Интересно рассмотреть во сколько раз увеличивается расчётная вероятность встречи двух одинаковых рецессивных гетерозиготных генов при направленном близкородственном скрещивании близких родственников представителя популяции взятого произвольно из большой популяции (инцест), по сравнению с расчётной вероятностью «встречи» 2-х генов, эмитирующих встречи в больших природных популяциях. И мы видим во сколько раз увеличивается вероятность встречи в случае близкородственного скрещивания по сравнению с процессом, проходящим в естественных условиях в большой популяции. Так, для очень многих аутосомных рецессивных наследственных заболеваний, которые, безусловно, находятся в «подвале» рецессивных генов (в связи с их большой вредностью), частота проявлений в большой популяции составляет 1 из 1000 до 1 из 200000. Эти значения выделены в таблице 2 жирным шрифтом. Вероятность их проявления в случае близкородственного скрещивания людей из этих естественных популяций увеличивается в от 32 до 448 раз по сравнению с произвольными встречами представителей этой популяции в естественных условиях, исключающих близкородственное скрещивание. Для единичных (первичных) мутаций в популяции эта вероятность увеличивается в ещё большее число раз – в 1000 раз (первая графа таблицы 2). Вероятность для «встречи» двух рецессивных генов при близкородственном скрещивании определяется, прежде всего, вероятностью попадания обладателя «нового» рецессивного гена в группу подвергнутых близкородственному скрещиванию. Эти данные показывают на сколько близкородственное скрещивание делает реальным для единичных рецессивных генов набрать необходимое количество для вхождения в гомозиготное состояние и включения в конкурентную борьбу в естественном отборе. Без близкородственного скрещивания вероятность завоевания рецессивным геном сколько ни будь значительной доли в популяции невозможна.
При этом рассуждении предполагается, что влияние других факторов на вероятность встречи двух одинаковых генов, как-то наличие и влияние доминантных генов в популяции, рождение одного из четырёх детей [3] с генами в гомозиготном состоянии, смертность зародыша до рождения (хотя «встреча» гетерозиготных генов была), и других факторов, будет одинаково влиять в обоих случаях независимо от размера популяции и в случае если «встреча» гомозиготной пары генов произошла при участии близких родственников. Учитывая это, мы можем значительно упростить расчёты и нагляднее показать работу собственно рецессивных генов без ожидания получения ошибки способной повлиять на конечный результат.
Таким образом, зная реальную статистику - вероятность рождения больных детей в естественных условиях большой популяции мы можем прикинуть вероятность рождения больного ребёнка при близкородственном скрещивании. 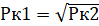 . Практическое применение данного вывода могло бы заключаться в определении вероятности рождения больного ребёнка при инцесте – крайней жёсткой форме близкородственного скрещивания. К сожалению, у меня нет данных подтверждающих или опровергающих данное рассуждение, однако у специалистов-медиков такие данные должны быть. Дополнительно замечу, что следует ожидать, в большой популяции и с «недостаточной однородностью» распространения рецессивных генов, что средняя оценка в большинстве случаях (на большинстве территорий) будет искажать истинное состояние в сторону завышения истиной концентрации в популяции. Значит, расчётная величина проявления рецессивного гена при близкородственном скрещивании для большинства территорий будет несколько завышена, а для отдельных территорий, где имеет место наибольшее местное распространение рецессивного гена - занижена. При этом, чем меньше величина вероятности проявления наследственного заболевания, тем больше ошибка в прогнозируемом близкородственном скрещивании причём в завышенную сторону (фактически ниже).
. Практическое применение данного вывода могло бы заключаться в определении вероятности рождения больного ребёнка при инцесте – крайней жёсткой форме близкородственного скрещивания. К сожалению, у меня нет данных подтверждающих или опровергающих данное рассуждение, однако у специалистов-медиков такие данные должны быть. Дополнительно замечу, что следует ожидать, в большой популяции и с «недостаточной однородностью» распространения рецессивных генов, что средняя оценка в большинстве случаях (на большинстве территорий) будет искажать истинное состояние в сторону завышения истиной концентрации в популяции. Значит, расчётная величина проявления рецессивного гена при близкородственном скрещивании для большинства территорий будет несколько завышена, а для отдельных территорий, где имеет место наибольшее местное распространение рецессивного гена - занижена. При этом, чем меньше величина вероятности проявления наследственного заболевания, тем больше ошибка в прогнозируемом близкородственном скрещивании причём в завышенную сторону (фактически ниже).
7. Но ведь в «подвале» находятся не только отрицательные рецессивные гены, но и положительные. Никакой из успешных генов не миновал в прошлом этого состояния, каким бы эффективным он ни был. Так же проявляют себя и положительные мутации. Мы можем перечислить целый ряд редких положительных (на наш взгляд) проявлений наследственности, вызванных встречей рецессивных генов в гомозиготном состоянии: это уже известные нам мутации гена ССR5Δ32 способные противостоять вирусу ВИЧ, мутация, обеспечивающая повышенную прочность костей, меньшую потребность во сне и отдыхе, способность зрения различать больше цветов, устойчивость к холоду и пониженному содержанию кислорода в воздухе. Проявлениями подобных мутаций можно объяснить и различные интеллектуальные способности, и рождение исключительно способных людей.
8. До сих пор мы рассматривали гены как самостоятельные, определяющие своё исключительное положение в «очереди» банка рецессивных генов и занять достойное место среди внедрённых доминирующих в популяции генов. Но есть и ещё один способ «незаслуженного» перемещения генов в ряду продвижения к увеличению численности и наоборот. В некотором смысле «по блату» или «за компанию», то есть, в связи с «родством» с положительным или отрицательным рецессивными генами. Понятно, что дети получают от родителей два комплекта генов: один от папы, другой от мамы. Они, в свою очередь, аналогично передают комплекты генов своим детям и так далее. Гены, обладающие способностью увеличивать численность своих обладателей увеличивают и численность своих «собратьев», доставшихся им от родителей и наоборот. Одна надежда, что со временем естественный отбор (если он есть) наведёт в этом порядок и каждый ген займёт своё место в соответствии со своими качествами.
9. Справедливости ради надо сказать несколько слов и о аутосомно-доминантных генах и их мутациях в том числе вызывающих наследственные заболевания. По логике вещей мутированных доминантных генов, вызывающих наследственные заболевания не должно быть, ведь они не могут «спрятаться» от естественного отбора в «подвале» и будут подвергнуты жестокому уничтожению если они будут вызывать сокращение численности потомства. Видимо, не все, а скорее даже большинство аутосомно-доминантных наследственных заболеваний так сильно не влияют на численность популяции и продолжают сопровождать нас и передаваться нашим потомкам. Их, как мне представляется, правильнее называть отклонениями или нарушениями наследственности, но не заболеваниями. В то же время, и множество незначительных отклонений может свести на нет наследственность популяции. Хотя нет. Как только эти недостатки будут серьёзно влиять на жизнеспособность и рождаемость естественный отбор с ними разберётся. Кроме того, видимо, «новые» рецессивные гены, обладающие лучшими качествами способны заменить доминантные гены в том числе и дефективные, если, конечно, будет работать естественный отбор.
2. «Скромные» рецессивные гены.
В нашем понимании, раз рецессивный ген положителен то он должен в итоге добиться своего однозначного доминирования в популяции. Например, ген, обеспечивающий противостояние вирусу, который способен уничтожить популяцию. Казалось бы, естественный отбор должен в первую очередь обеспечить абсолютное доминирование данного гена. С другой стороны, вирус начинает угрожать популяции только тогда, когда возникает эпидемия. В период, когда нет эпидемии обладатели спасительного гена не имеют постоянного преимущества перед не обладающими данным геном. Более того, как мне представляется, спасительный ген совсем не обязан в отсутствии агрессии со стороны вируса быть хотя бы нейтральным. Ведь он заменяет ген, выбранный отбором в условиях отсутствия отрицательного влияния вируса. В условиях смертельной опасности, когда прочие отрицательные проявления спасительного гена не могли сыграть решающей роли, он будет увеличивать своё представительство в популяции. Однако, вероятнее всего, он будет иметь побочный отрицательный эффект, как все лекарства, и обладатели его будут уничтожаться естественным отбором при отсутствии эпидемии до тех пор, пока не возникнет новая эпидемия или число обладателей «спасительного» гена уменьшится до критической величины, когда можно спрятаться в «подвале». Таким образом, концентрация спасительного гена в гомозиготном состоянии будет изменяться в определённых пределах. Всё зависит от способности вируса создавать эпидемические ситуации – частоты возникновения эпидемий, степени опасности вируса и вредности гена в условиях отсутствия эпидемии. Такие спасительные гены я бы назвал «генами-санитарами», но когда «санитаров» в популяции неоправданно много, это приводит к ухудшению жизнеспособности популяции, а возникает их много, как, впрочем, и других малочисленных генов в период близкородственного скрещивания. Но и потеря спасительного гена может привести к непоправимой потере. Наверняка, существуют и другие гены способные, с одной стороны, оказывать положительное влияние на состояние популяции в одних условиях, с другой - отрицательное в других. За счёт естественного отбора соотношением их в популяции позволяет получать приемлемый, может быть не идеальный, но приемлемый вариант в зависимости от условий. Какое-то время не используемые гены в случае если их малой концентрация в популяции будут храниться в «подвале» - недоступном для естественного отбора. В принципе, это мы можем наблюдать на примере рецессивного гена ССR5Δ32, способного противостоять ВИЧ/СПИД вирусу[5]. Я думаю, что когда-то рецессивный ген был и у африканцев, но был утрачен ими в связи с длительным отсутствием эпидемий подобных вирусов. Ведь вирусы не сегодня родились, хотя и таких больших людских популяций, непосредственно связанных между собой как в последние тысячи лет до сих пор не было. А ведь это благотворная среда для возникновения эпидемий. В тоже время, вирусы где-то у кого-то жили и размножались и до возникновения больших перемешиваемых популяций.
Ещё один случай. Рассмотрим абстрактный пример: допустим, что существует ген, способствующий устойчивости организма к пониженным температурам. А почему нет? Тогда в условиях с холодным климатом естественный отбор будет увеличивать обладателей таких генов в популяции. Однако, как правило, улучшение одного вызывает нарушение и ухудшение другого. В этих условиях с мягким температурным режимом он же – естественный отбор - будет уменьшать обладателей данного гена. Значит, где-то, по середине крайних температурных режимов возникнет оптимальное равновесие рецессивного гена и другого его варианта, применительно к конкретным условиям. Любой сдвиг фактической средней температуры от оптимального значения будет вызывать ухудшение работы системы и со временем будет стремиться изменять соотношение обладателей разных генов. Такие гены в общем виде можно назвать «генами – балансёрами» (или генами компенсаторами). Если рецессивные гены из «подвала» способны только в результате близкородственного скрещивания попадать в число работающих – гомозиготных, то «балансирующие гены» без близкородственного скрещивания способны участвовать в естественном отборе. В аналогичной ситуации находятся и другие гены набирающие или наоборот теряющие своё представительство в популяции.
«Генов – балансёров», видимо, множество. Можно предположить, что и гены, обеспечивающие усвоение кислород из атмосферы относятся к этой категории генов. В популяциях, проживающих в условиях высокогорья, где атмосфера разряжена и тот же объём воздуха содержит меньше кислорода в местных популяциях наблюдаются генетические изменения. Интересно и то, что на разных высокогорных территориях на Земле одно и тоже качество – усвоение организмом достаточного количества кислорода в условиях разрежённости воздуха - обеспечивается мутациями различных генов[6].
В заглавии этого раздела я назвал подобные гены «скромными» по той причине, что они не стремятся к полному доминированию в популяции и сосуществуют на равне с другими.
3. Факторы, влияющие на наличие и разнообразие рецессивных генов в популяции и их сохранение.
Мы уже давно определили важность разнообразия рецессивных генов в популяции. Осталось только уточнить, что всё разнообразие людей на планете определяется, прежде всего, разнообразием генов и рецессивных в первую очередь, поскольку, именно они разные во всех популяциях и именно они определяют последние изменения наследственности, а значит и формирование отличий различных представителей внутри популяции.
Но что же, в прошлой жизни популяции, влияло и влияет сейчас на это разнообразие?
1) Рождаемость. Теоретически, для передачи детям от родителей полного пакета ДНК необходимо иметь потомство не менее 4-х детей[3]. Это при равномерном распределении генов родителей среди детей, что, конечно же, практически не возможно. Для большей уверенности в передачи всех генов детям необходимо ещё больше детей. Наибольшей опасности потери при сокращении рождаемости подвергаются гены, представленные в популяции минимальным представительством.
2) Ранее мы говорили, что вероятнее всего мутационные штурмы проходят в изолированной части большой популяции, пример – евреи-ашкенази. Однако, именно изолированная часть содержит только часть возможного ассортимента рецессивных генов популяции. Чем меньше часть популяции, участвующей в мутационном штурме - тем меньшая часть от общего банка рецессивных генов будет участвовать в процессе отбора, и меньшая доля рецессивных генов будет передана новой популяции после мутационного штурма и, вероятно, с меньшим разнообразием. В процессе образовании «бутылочного горлышка», при отрицательном воздействием внешней среды, происходит значительное сокращение численности и разнообразия рецессивных генов. Только небольшая часть обладателей спасительного гена проходит через «бутылочное горлышко» и «проносят» с собой только часть разнообразия рецессивных генов изолированной части популяции.
3) В процессе застоя происходит естественный отбор и обладатели устаревших генов сокращают свою долю. В то же время некоторой гарантией сохранности малочисленных генов является «подвал», гарантирующий сохранность малочисленных рецессивных генов.
4) Увеличение разнообразия рецессивных генов малой популяции, прошедшей «бутылочное горлышко» и мутационный штурм, происходит при слиянии её с частью большой популяции. Правильнее сказать, что это не увеличение разнообразия, а лишь частичная компенсация потерь разнообразия рецессивных генов в результате мутационного штурма. Отсюда следует вывод: чем больше мутационных штурмов прошла популяция, тем меньше разнообразие рецессивных генов сохраняется в банке популяции и наоборот, чем больше разнообразие рецессивных генов, тем меньше мутационных штурмов прошла популяция. Одновременно стоит отметить, что в процессе близкородственного скрещивания возможно увеличение численности рецессивных генов в том числе и малочисленных, участвующих в близкородственном скрещивании, что способствует сохранению генов. Однако, это не увеличивает разнообразия генов.
5) Позже, мы увидим, что при смешении генетически неродственных популяций у потомства возможны потери рецессивных генов как в гомозиготном состоянии, так и в гетерозиготном состояниях и, как следствие, потери работающих положительных качеств присущих каждой из популяций в отдельности, выработанных в популяциях после разделения общей популяции на самостоятельные и уменьшение концентрации положительных гетерозиготных генов. Проявления этого процесса при разведении животных получило название аутбредной депрессии. Подробней этот процесс мы рассмотрим ниже, а сейчас важно уяснить, что это не увеличение разнообразия рецессивных генов двух популяций, хотя общая популяция будет иметь большее разнообразие рецессивных генов, чем каждая предшествующая в отдельности. При этом будет иметь место перевод генов из гомозиготного состояния, то есть, рабочего, в гетерозиготное – спящие и сокращение доли гетерозиготных генов в общей популяции за счёт «разбавления» генами другой популяции.
6) Рассматривая процессы, протекающие в той или иной популяции мы видим, что в процессе эволюции меняется количество (доля) одних рецессивных генов – обладателей положительных генов должно увеличиваться, а, следовательно, уменьшается количество (доля) обладателей других.
7) Рецессивные гены, представленные небольшим количеством, имеют большую вероятность быть потерянными. Об этом я бездоказательно ранее говорил неоднократно. Это вытекает из рассмотрения вероятности событий передачи генов следующему поколению и потери генов при этом. И не только. В любом процессе, определяемом как вероятностное событие больше вероятность потерять малочисленно представленные гены. Рассматривая популяцию и малое представительство в ней того или иного гена мы имеем дело с вероятностными событиями при малой численности одинаковых рецессивных генов. При малом числе событий вероятность отклонений от средней статистической величины тем больше чем меньше число событий – тем больше величина флуктуации. Флуктуация – любое случайное отклонение какой-либо величины. Отклонения от этой величины может быть, как в большую, так и в меньшую стороны – колеблется вокруг средней величины. Однако, отклонение в меньшую грозит исчезновением редкого рецессивного гена и обратного пути уже не будет. Он будет потерян навсегда сначала в какой-то части популяции, а затем и для всей. Частично отражение этого процесса нашло в правиле, сформулированном получившим название «эффект основателя» – основателя популяции[9]. Он был описан Эрнстом Майром в 1942году. После отделения малой популяции от большой вероятно нарушение соотношения долей обладателей тех или иных генов. Хотя, «эффект основателя», вернее, наблюдаемое изменение наследственности, включает и другие механизмы изменения доли генов в новой популяции в том числе и за счёт естественного отбора и случайных факторов, поскольку он в общем виде не рассматривает природу явления, а лишь констатирует наблюдаемый факт. Очень вероятно и близкородственное скрещивание поскольку популяция маленькая. и это способствует проявлению совершенно новых генов. Об этом мы поговорим позже.
8. Окончательной потере генов, в том числе и отрицательных, в результате естественного отбора, противостоит «подвал», в котором рецессивные гены находятся в основном в гетерозиготном состоянии и не проявляют своих качеств в том числе отрицательных. Там может храниться множество редких генов в том числе очень старых, давно вышедших из «массовой эксплуатации». В определённых условиях они могут проявлять себя.
9. Все перечисленные выше факторы способны лишь уменьшить в той или иной степени разнообразие рецессивных генов. И только один из всех механизмов способных разнообразить банк рецессивных генов, более того предложить новые варианты генов — это естественные мутации рецессивных генов. Однако, как мы уже неоднократно повторяли, основная масса мутаций носит отрицательный характер. Что бы выйти из «подвала» банка рецессивных генов мутированный ген должен пройти не один мутационный штурм и получить определённую долю представительства за счёт естественного отбора. Именно такой многоступенчатый отбор и определяет наличие в «очереди» исключительно положительных генов в средней и верхних частях графика (см. рис. 1).
Возникает естественный вопрос: почему раньше происходил процесс накопления разнообразных и, прежде всего, положительных рецессивных генов в отличие от сегодняшнего этапа, в частности, человеком, когда мы наблюдаем потери накопленных ранее рецессивных генов? (Именно так мне представляется, хотя, могут быть и не согласные со мной. К рассмотрению баланса приход/расход мы ещё вернёмся). И, как ни странно, это будут те же механизмы, которые мы рассматривали ранее как уменьшающие разнообразие: мутационный штурм и естественный отбор, но в других условиях они будут работать иначе. А условия эти заключаются в изолированности отдельных небольших групп наших далёких предков, где процессы мутационного штурма проходили чаще из-за лёгкости достижения «бутылочного горлышка» и процесса близкородственного скрещивания. Раньше интенсивней проходил естественный отбор, формирование «очереди» положительных рецессивных генов и внедрение прогрессивных мутаций в связи с жесткими условиями жизни, когда каждое усовершенствование человеческого организма определяло его выживаемость. И, как это ни странно, интеллектуальные способности были более востребованы чем сегодня. Сейчас часто происходят процессы неродственного скрещивания, об отрицательных последствиях которого мы поговорим отдельно. Так что, сохранение разнообразия рецессивных генов в той или иной популяции носит первостепенное значение. Надеяться на отбор новых положительных генов при отсутствии естественного отбора (о чём мы поговорим позже) не представляется возможным.
«Старые» положительные завоевавшие большинство в гомозиготном состоянии могут работать долго даже в условиях отсутствия естественного отбора, однако, потеряв их в условиях отсутствия естественного отбора восстановить их будет невозможно.
Тот или иной банк рецессивных генов популяции как совокупность множества банков отдельных представителей популяции — это продукт эволюционного развития популяции, вида, есть буквально единственный самый главный продукт национального достояния популяции.
В итоге мы можем констатировать, что важнейшим элементом эволюционного развития является сохранение сегодняшнего разнообразия генов в популяциях как в гомозиготном, так и в гетерозиготном состояниях.
Наша задача по возможности уменьшать влияние перечисленных выше отрицательных факторов, влияющих на сокращение разнообразия генов в популяции. Мы теряем то, что уже никогда не сможем приобрести. Это тема для серьёзного разговора и серьёзных исследований.
4. Влияние размера популяции на ход мутационного штурма.
1. Теперь о популяции. В статье[4] мы рассмотрели популяцию как генетически однородную структуру, где все члены популяции перемешаны до абсолютного однообразия. Реально большая популяция не однородна. И каждый член популяции обладает своим исключительным набором не только работающих генов, но в ещё большей степени различным набором рецессивных генов в гетерозиготном состоянии. Это и понятно, ведь работающие гены уже прошли отсев естественным отбором, в то время как рецессивные гетерозиготные не прошли полную фильтрацию и отсев по эффективности, а в «подвале» находится исключительное разнообразие рецессивных генов, накопленное за миллионы лет и у каждого оно своё.
2. Наглядно популяцию можно представить, как луг с определённым разнообразием представленных на каждом квадратном метре различных растений. Часть растений, ну, например, одуванчик или пырей, какой ни будь (прошу прощения у ботаников за бесцеремонное использование в качестве макета их любимцев), будут представлены на каждом квадратном метре нашего луга. Во многих местах мы увидим, возможно, подорожник, клевер, гвоздики, ромашки, колокольчики. Отдельными островками будут представлены те же цветы, но не отдельными кустиками, а целыми колониями. Единично представлены в отдельных местах «иностранцы» - мак, дикие лилии или иван-чай, занесённые сюда недавно и не нашедшие ещё массового распространения. Все растения борются за занятие территории и разбрасывают свои семена повсюду, но везде они будут представлены разным количеством – где-то больше, где-то меньше, а где-то совсем не представлены. Вот так можно единовременно представить картину распространения рецессивных генов в популяции. Разнообразная картина, но всех их объединяет некое единство – много одинаковых растений, если их сравнивать с растительностью, ну скажем, саванн в Африке. Это в нашем сравнении будет другая популяция. В популяции людей с генетической точки зрения всё очень похоже. В этом мы можем убедиться вспомнив распространение рецессивного мутированного гена ССR5Δ32 в Центральной Европе, когда его концентрация в гетерозиготном состоянии составляла от 5% до 15% [7], с градиентом увеличения концентрации с юга на север, а у архангельских поморов составила даже 30%[8]. Чем Вам не какой-нибудь колокольчик на европейской лужайке. И в Африке этот ген тоже не представлен. Как в человеческих популяциях, так и среди растений по труднодоступным окраинам, куда проникновение хорошо «перемешиваемой» части популяции затруднено, мы можем наблюдать более генетически однородные части популяции. То есть, в этом случае на лугу по окраинам мы могли бы наблюдать большее скопление одинаковых цветов, а, следовательно, там чаще происходят близкородственные скрещивания и даже свои микро мутационные штурмы и естественный отбор. Здесь складываются свои микро популяции, отличающиеся доминированием своего «нового» рецессивного гена в гомозиготном состоянии. Естественно, что на разных окраинах большой популяции мы будем видеть аналогичные картины и преобладание различных рецессивных генов. Причём, это касается не только рецессивных генов в борьбе с доминантными, но и с ранее завоевавшими большинство другими рецессивными генами. Доминантные гены будут так же бороться между собой и с рецессивными. Этот процесс будет носить ещё более ярко выраженный характер при полной изоляции микро популяций от большой популяции. Нечто подобное как снижение и смещение генетического разнообразия при заселении малым количеством представителей рассматриваемого вида новой географической территории было замечено Эрнстем Майром[9]. Он назвал это явление «эффект основателя». Он считал, что в малых популяциях разные гены могут завоевать большинство в зависимости от того, кто при формировании новой малой популяции был «основателем». Конечно же, «эффект основателя» играет определённую роль в формировании новых малых популяций, но не исчерпывается только этим механизмом не является объективным объяснением различной наследственности на изолированных территориях. Он не отражает полной картины механизма формирования малых популяций, а лишь выделяет одну из составляющих. Хотя, факт наличия различий в разных малых популяциях по отношению к большим «материнским» отражён правильно.
Реальная популяция не однородна. Наибольшие отклонения от среднего состояния можно наблюдать в частично изолированных краях популяции и особенно в изолированных микро популяциях, отделившихся от основной. Именно там более интенсивно проходит близкородственное скрещивание и естественный отбор по набравшим достаточное большинство обладателей рецессивных генов в гомозиготном состоянии меняет характерные для популяции качества. Там чаще происходит микро мутационный штурм и естественный отбор с завоеванием большинства обладателями «нового» положительного гена. Аналогичную ситуацию мы получаем в случае возникновения «бутылочного горлышка» - один из случаев микро популяции. «Проходящую» через «бутылочное горлышко» популяцию нужно рассматривать как микро популяцию. «Бутылочное горлышко», как правило, возникает в результате какого-либо отрицательного воздействия на популяцию. Что приводит к сокращению её численности.
Отдельно стоит упомянуть «единичные» популяции – популяции, возникшие в результате близкородственного скрещивания нескольких единиц представителей популяции. Это даже не микро популяции прохождения через «бутылочное горлышко». Мутационный штурм в этих условиях способен пробудить самые единичные рецессивные гены, представленные в «подвале», что чревато радикальными изменениями в формировании нового вида. Примером таких популяций могут быть островные популяции животных, переселившиеся туда с материка. Поскольку в подобных процессах мы можем наблюдать разнообразие нескольких процессов, происходящих при близкородственном скрещивании мы рассмотрим всё это при рассмотрении различных вариантов мутационного штурма в одной из следующих статей.
5. О роли теории мутационного штурма и естественного отбора как части общей картины эволюционного процесса.
Исчерпывает ли теория мутационного штурма и естественного отбора в полной мере эволюционный процесс? Конечно же, нет. И первое, что сразу говорит о ограниченности теории — это факт самого наличия рецессивных и доминантных генов. А именно: Понятно, что все мы мутанты. То есть, все сегодняшние наши гены есть мутированные рецессивные, «завоевавшие» большинство обладателей. Но фактически, по их «поведению», мы наблюдаем, что среди них есть как доминантные, так и рецессивные гены, а это значит, что в силу каких-то причин одни, в прошлом рецессивные гены, проявляют по отношению к другим доминантные качества, что никак не учитывает теория мутационного штурма и естественного отбора. Кроме того, опыты Менделя и дальнейшие исследования показывали, что спящие гены в гетерозиготном состоянии не работают, хотя есть подтверждения, что имеет место и влияния рецессивных генов в гетерозиготном состоянии на наследственность. Более того, обнаружен, и даже используется сегодня, для выведения новых сортов растений (так называемых, генно-модифицированных растений), перенос генов «по горизонтали» от одного биологического организма другому – от представителя одного вида другому. И в естественных природных условиях возможны такие случаи, когда работали подобные механизмы, однако это не носит массового характера, а скорее исключение. В противном случае мы все бы были различными: кто-то с хоботом, кто-то с шеей как у жирафа, кто-то с хвостом как у павлина. Природа многообразна и ещё множество законов даже в популяционной генетике должны ещё быть поняты.
Подобное заявление необходимо для понимания роли мутационного штурма и естественного отбора в эволюционном процессе. Это не есть заключительное заявление, подводящие итоги данной работы, нет. В следующих статьях мы рассмотрим, обещанные ранее, варианты мутационного штурма и специфики естественного отбора в современных условиях в том числе применительно к человеку. А также вопросы возможной деградации человечества как вида.
- Занегин М. А. О роли рецессивных генов в эволюционном процессе // Научные высказывания. 2023. №2 (26). С. 13-19. URL: https://nvjournal.ru/article/O_roli_retsessivnyh_genov_v_evoljutsionnom_protsesse
- Занегин М. А. Эволюционный процесс. Этапы «Мутационного штурма и естественного отбора» // Научные высказывания. 2023. №4 (28). URL: https://nvjournal.ru/article/Evoljutsionnyj_protsess_Etapy_Mutatsionnogo_shturma_i_estestvennogo_otbora
- Занегин М. А. Особенности состояния ДНК в субэтнической группе евреев-ашкенази. «Мутационный штурм» и «эволюционная пирамида» // Научные высказывания. 2023. №5 (29). С. 25-30. URL: https://nvjournal.ru/article/Osobennosti_sostojanija_DNK_v_subetnicheskoj_gruppe_evreev-ashkenazi_Mutatsionnyj_shturm_i_evoljutsionnaja_piramida
- Занегин М.А. Как теория мутационного штурма и естественного отбора дополняет и развивает дарвиновскую теорию https://nvjournal.ru/article/Kak_teorija_mutatsionnogo_shturma_i_estestvennogo_otbora_dopolnjaet_i_razvivaet_darvinovskuju_teoriju
- Занегин М. А. Примеры эволюционного развития, подтверждающие правильность теории мутационного штурма и естественного отбора. https://nvjournal.ru/article/Primery_evoljutsionnogo_razvitija_podtverzhdajuschie_pravilnost_teorii_mutatsionnogo_shturma_i_estestvennogo_otbora
- Центр палеоэтнологических исследований. А. С. Коньков Генетические адаптации человека к условиям высокогорья https://paleocentrum.ru/science/geneticheskie-adaptatsii-cheloveka-k-usloviyam-vysokogorya.html
- Pardis C. Sabeti, Emily Walsh, Steve F. Schaffner, Patrick Varilly, Ben Fry, Holli B. Hutcheson, Mike Cullen, Tarjei S. Mikkelsen, Jessica Roy, Nick Patterson, Richard Cooper, David Reich, David Altshuler, Stephen O’Brien, Eric S. Lander. The case for selection at CCR5-Delta32 (англ.) // PLoS Biology. — 2005. — Vol. 3, no. 11. — P. e378. — doi:10.1371/journal.pbio.0030378. — PMID 16248677.
- Кофиади И. А. Генетическая устойчивость к заражению ВИЧ и развитию СПИД в популяциях России и сопредельных государств: Автореферат. — Москва, 2008. Архивировано 23 ноября 2011 года.
- Провайн, В. Б. (2004). "Эрнст Майр: генетика и видообразование". Генетика. 167 (3): 1041-6. doi:10.1093/генетика /167.3.1041. PMC 1470966. PMID 15280221.
Научные высказывания #99