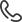Физический взгляд на борьбу растений со стрессом
В этой статье мы хотим вскрыть вопросы:
- Что же первично в условиях мультистресса? Успешная борьба с недостатком полива, как самый древнейший и важнейший для существования растения механизм, запускает цепочку борьбы за жизнь и с другими факторами стресса или же защитный ответ растений формируется на каждый стрессовый фактор сугубо специфически?
- Склонны ли растения рисковать и предпочитать обильный по объёму, но редкий во времени полив, минимальному по объёму, но регулярному во времени поливу?
- Подчиняется ли борьба растений со стрессом какому-либо физическому закону и может ли быть рассчитана и отражена математически, в цифровом эквиваленте, т.к. по этому поводу в биологии фигурируют субъективные формулировки: относительно засухоустойчивое растение, сравнительно засухоустойчивое растение , достаточно засухоустойчивое растение, что глубинно не раскрывает процесс борьбы со стрессом.

Засуха, засоление и тяжелые металлы наиболее распространенные стрессовые факторы, негативно влияющие на рост и развитие растений. Растения на эти стрессовые факторы реагируют на молекулярном, клеточном и физиологическом уровнях через сложную сеть восприятия и передачи сигналов стресса, которая впоследствии инициирует множество защитных механизмов. Регуляторы важнейших абиотических стрессовых реакций записаны в геном растения, запускающий активацию защитной цепочки преобразований, отвечающих за борьбу и выживаемость [1,с.6-8]. Наиболее древнейшим абиотическим фактором, с которым сталкивались и продолжают сталкиваться растения, является засуха. Данный стрессовый фактор на первых позициях и в условиях современного мира с происходящими процессами глобального потепления, изменения климата на более засушливый и истощения ресурса пресной воды, а 70% мировых земель признаны весьма условно пригодными для земледелия по совокупности неблагоприятных факторов: засухи или, наоборот, гипотермии, эрозии, засоления и загрязнения тяжёлыми металлами, т.е. представляют собой для растений условия мультистресса.
Растения развили способность адаптироваться к суровым условиям окружающей среды, причём в природе современного мира чаще всего наблюдается комплексное воздействие стрессовых факторов. Значит ли это, что растения, которые могут пережить один стресс, окажутся более приспособленными ко второму? Третьему? Одновременно?
Безусловно, все растения обладают способностью бороться с засухой, но сильно различаются по данному параметру.
Является ли ключевым моментом запуска защитных систем для растений фактор засухи (недостаточного полива), как наиболее древний абиотический фактор, известный растениям, ведь способность успешно его преодолевать кодируется экспрессией сотен-тысяч защитных генов? Может в этой длинной цепочке активации защитных генов от недостаточного полива иметь место одновременная активация защиты от других стрессовых факторов? Или же для каждого стрессового фактора запускается своя, уникальная последовательность экспрессии генов?
В этой работе мы хотим вскрыть вопросы:
- Что же первично в условиях мультистресса? Успешная борьба с недостатком полива, как самый древнейший и важнейший для существования растения механизм, запускает цепочку борьбы за жизнь и с другими факторами стресса или же защитный ответ растений формируется на каждый стрессовый фактор сугубо специфически?
- Склонны ли растения рисковать и предпочитать обильный по объёму, но редкий во времени полив, минимальному по объёму, но регулярному во времени поливу?
- Подчиняется ли борьба растений со стрессом какому-либо физическому закону и может ли быть рассчитана и отражена математически, в цифровом эквиваленте, т.к. по этому поводу в биологии фигурируют субъективные формулировки: относительно засухоустойчивое растение, сравнительно засухоустойчивое растение , достаточно засухоустойчивое растение, что глубинно не раскрывает процесс борьбы со стрессом.
Здесь следует упомянуть, что всё же есть лабораторно-полевой способ расчёта засухоустойчивости растений по уровню оценки оводнённости тканей, по определению водного дефицита, определению водоудерживающей способности листьев, однако для этих методов требуется сожжение в термостате точно измеряной навески листьев при высоких температурах, плюс—возможность задать стабильный параметр влажности на протяжении всего исследования [2,с.43-46]. Громоздкий метод для простого обывателя и строго специфический именно для оценки засухоустойчивости. А как же тогда рассчитать устойчивость к другим видам стресса?
Если в физике найдётся эквивалент закона, к которому можно адаптировать борьбу растений со стрессом, в т.ч. с недостаточным уровнем полива—это значительно упростит процесс оценки стрессоустойчивости по каждому фактору и сможет привести субъективные формулировки к реальному цифровому выражению этих понятий.
Ход работы: высаживание нескольких ростков каждого вида растения в условия с разделением корневой системы каждого ростка на два компонента, в изолированные друг от друга ёмкости, с различным режимом полива в каждой, однако, единой вегетативной частью. Опыт дублируется в чернозёме и обеднённой, засоленной почве с примесью тяжёлых металлов (рис.1).
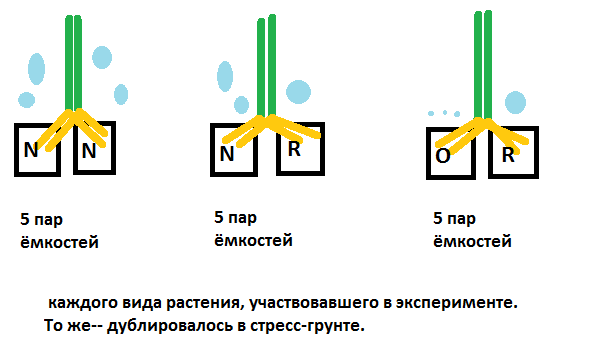
Рис.1. Наглядная методология посадки и различного вида полива растений в эксперименте с чернозёмом и стресс-почвой.
1 фаза эксперимента: укоренение/ проращивание в почве (чернозём) различных видов растений, различающихся между собой по способности противостоять засухе. (1 неделя).
2 фаза эксперимента: проверка склонности к риску различного типа полива—разделение в ёмкости с чернозёмом корневой системы на два изолированных друг от друга компонента. (укоренение—5 дней, различный режим полива—25 дней).
Всего 5 пар ёмкостей для растений одного вида, у которых разделена корневая система на два пучка. Каждая пара ёмкостей с растениями запланировано прекращала участие в эксперименте соответственно на 5-й, 10-й, 15-й, 20-й и 25-й день полива для взвешивания вегетативной и корневой частей.
3 фаза эксперимента (для тех видов растений, кто успешно преодолел 2 фазу, т.е. сохранил живую вегетативную часть в чернозёме на 25-й день полива): укоренение на обеднённой, засоленной, загрязнённой тяжёлыми металлами почве—новые ростки тех же видов растений (1,5 недели)
4 фаза эксперимента: разделение в ёмкости с обеднённой, засоленной почвой с примесью тяжёлых металлов, корневой системы на два изолированных друг от друга компонента с различным режимом полива ( 5 дней—укоренение, 25 дней—различный режим полива). По сути, это та же вторая часть эксперимента, но с новыми парами тех же видов растений в ещё более суровых условиях—засоления, обеднения почвы, примеси тяжёлых металлов.
Как нами создавалась обеднённая засоленная почва с примесью тяжёлых металлов (рис.2-5)?
В равных частях бралась эрозированная почва со склона в непосредственной близости от железной дороги, что также указывает на то, что в её составе имелась примесь тяжёлых металлов и чистый речной песок (кремнезём). Для большей уверенности в наличии тяжёлых металлов добавлена размолотая коррозированная металлическая стружка. Для защелачивания—вулканическая глина, размолотый силикатный кирпич, мел (известняк) до показателя рН на почвометре 8.
Однако, в данном виде почвы в чистом виде ни одно растение не выжило, поэтому данная часть эксперимента повторялась в почве, имеющей 1/3 указанного состава, 2/3 же объёма занимал чернозём. Данный тип получившейся почвы далее в нашей работе фигурирует под понятием стресс-почва.




Рис.2-5. Процесс создания стресс-почвы.
Единые режимы полива в эксперименте для каждого вида растения:
- Норма-норма.
- Норма- риск.
- Опрыскивание- риск.
За понятие нормы брался уровень увлажнённости почвы 5 по почвометру. ( 20 мл ежедневно в ёмкость объёмом 200 мл).
За понятие риск брался уровень увлажнённости почвы 5 по почвометру, с последующим снижением показателя до 0. ( 40 мл через каждые 3 дня).
За понятие опрыскивание брался уровень увлажнённости почвы 1 по почвометру (ежедневные опрыскивания почвы из пуливеризатора—5 нажатий).
Растения, изначально участвовавшие в эксперименте: горох, кукуруза, рожь, пшеница, полба (дикая пшеница), хлорофитум, помидоры, клевер. Однако, чтобы успешно разделить корневую систему на две розетки, пришлось отказаться от растений со стержневой моноподиальной корневой системой: гороха, помидоров, в пользу мочковатой корневой системы: рожь, пшеница, полба, кукуруза, клевер.
Растения, сохранившие живой вегетативную часть после 2 фазы и участвующие далее до полного окончания эксперимента : рожь, пшеница, кукуруза. Т.е. наиболее сильные претенденты на статус засухоустойчивых растений.
В каждую пару ёмкостей высаживалось по три растения для возможности более точного измерения массы корней и вегетативной части на ювелирных весах с погрешностью измерений данного прибора 0,01 г.
Эксперимент в ходу наглядно отображён на фотографиях (рис.6-7).


Рис. 6-7: Эксперимент в ходу.
Результаты работы:
- Кукуруза (корневая часть). Представлены рубрики с выжившей вегетативной (надземной) частью к концу эксперимента, т.е. на 25-й день полива.
Растения в грунте N/N и N/R, а также стресс-почве N/N погибли соответственно на 1-й, 2-й, 1-й неделе испытания (Таблица 1).
Таблица 1.
Убыль корневой массы кукурузы в ходе эксперимента.
|
Дни экспериментального полива |
Грунт O /R |
Стресс-грунт O/R |
Стресс-грунт N/R |
|
0 день |
0,68 |
0,68 |
0,68 |
|
5 день |
0,60 |
0,58 |
0,48 |
|
10 день |
0,53 |
0,49 |
0,32 |
|
15 день |
0,44 |
0,40 |
0,22 |
|
20 день |
0,38 |
0,35 |
0,16 |
|
25 день |
Ʃ 0,35
O /R 0,14/0,21
|
Ʃ 0,31
O /R 0,17/0,14
|
Ʃ 0,14
N/R 0,08/0,06
|
График зависимости убыли корневой массы кукурузы (г)—Оу от времени (дни)—Ох.
За исходную массу корневой системы (день 1) бралась масса корней трёх здоровых пророщенных/укоренившихся растений после 1-й фазы испытаний. Для кукурузы это 0,68 г. Извлеченные из земли корни промывались и высушивались на ватных дисках естественным путём 1 сутки, только затем взвешивались.
Таблица 2.
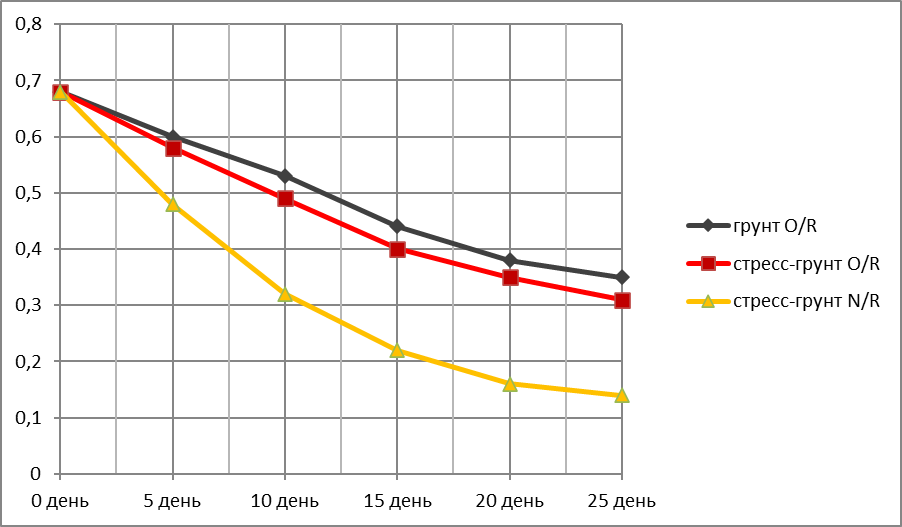
Убыль массы вегетативной части кукурузы в ходе эксперимента.
Представлены образцы, сохранившие живой вегетативную часть все 25 дней опыта.
Растения в грунте N/N и N/R и стресс-почве N/N погибли соответственно на 1-й,2-й и 1-й неделе испытания.
|
Дни экспериментального полива |
Грунт O /R |
Стресс-грунт O/R |
Стресс-грунт N/R |
|
0 день |
5,56 |
5,56 |
5,56 |
|
5 день |
4,87 |
4,65 |
4,26 |
|
10 день |
4,18 |
4,05 |
3,23 |
|
15 день |
3,66 |
3,54 |
2,56 |
|
20 день |
3,24 |
3,05 |
2,02 |
|
25 день |
3,05
|
2,82
|
1,7
|
График зависимости убыли массы надземной (вегетативной) части кукурузы (г)—Оу от времени (дни)—Ох.
За исходную массу вегетативной части (день 1) бралась масса трёх здоровых средних по размеру пророщенных растений в чернозёме без стресса разделения корней после 1-й фазы испытаний. Для кукурузы это 5,56 г (Таблица 2).
- Рожь (корневая часть). Представлены рубрики с выжившей вегетативной (надземной) частью к концу эксперимента, т.е. на 25-й день полива.
Растения в грунте N/N и N/R, а также стресс-почве N/N и стресс-почве N/R погибли соответственно на 1-й, 2-й, 1-й и 1-й неделе испытания.
Таблица 3.
Убыль корневой массы ржи в ходе эксперимента.
|
Дни экспериментального полива |
Грунт O /R |
Стресс-грунт O/R |
|
0 день |
0,36 |
0,36 |
|
5 день |
0,28 |
0,23 |
|
10 день |
0,19 |
0,15 |
|
15 день |
0,11 |
0,08 |
|
20 день |
0,07 |
0,03 |
|
25 день |
Ʃ 0.05
O /R 0,03/0,02 |
Ʃ 0,01
O /R ? |
График зависимости убыли корневой массы ржи (г)—Оу от времени (дни)—Ох.
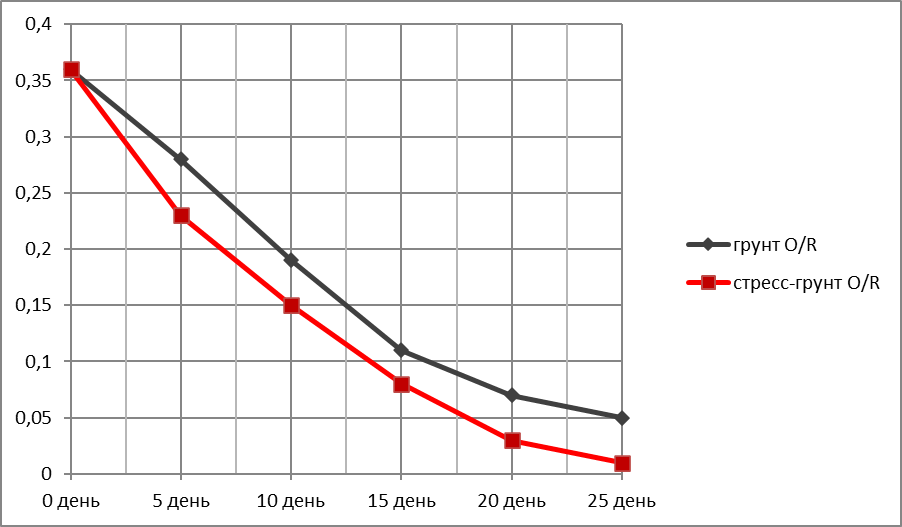
За исходную массу корневой системы (день 1) бралась масса корней трёх здоровых пророщенных/укоренившихся растений после 1-й фазы испытаний. Для ржи это 0,36 г (Таблица 3).
Взвешивание растений с помощью высокоточных весов представлено на фотографиях (рис.8-9).


Рис. 8-9: взвешивание корневой и вегетативной массы одного здорового растения ржи с помощью высокоточных весов с ценой деления 0,01г наглядно.
Таблица 4.
Убыль массы вегетативной части ржи в ходе эксперимента.
Представлены образцы, сохранившие живой вегетативную часть все 25 дней опыта.
Растения в грунте N/N и N/R и стресс-почве N/N и стресс-почве N/R погибли соответственно на 1-й и 2-й ; 1-й и 1-й неделе испытания.
|
Дни экспериментального полива |
Грунт O /R |
Стресс-грунт O/R |
|
0 день |
0,69 |
0,69 |
|
5 день |
0,56 |
0,5 |
|
10 день |
0,43 |
0,36 |
|
15 день |
0,32 |
0,25 |
|
20 день |
0,23 |
0,15 |
|
25 день |
0,15
|
0,07
|
График зависимости убыли массы надземной (вегетативной) части ржи (г)—Оу от времени (дни)—Ох.
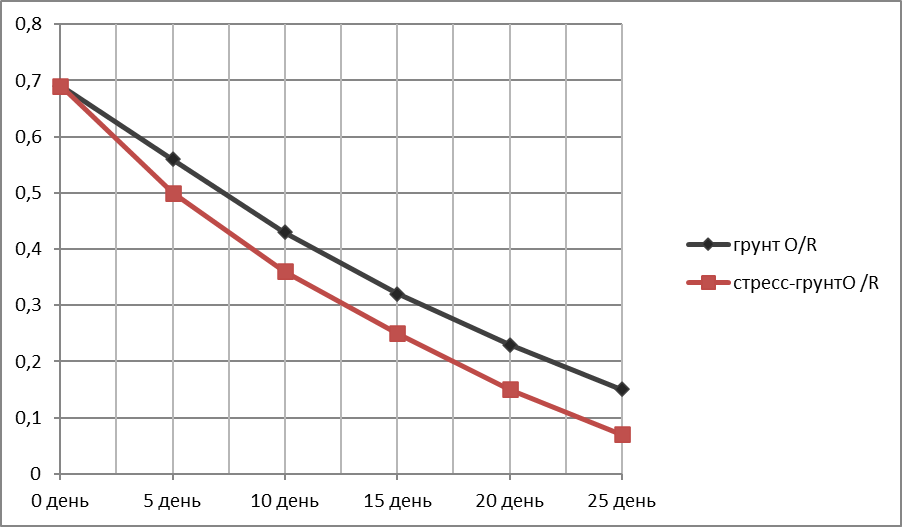
За исходную массу вегетативной части (день 1) бралась масса трёх здоровых средних по размеру пророщенных растений в чернозёме без стресса разделения корней после 1-й фазы испытаний. Для ржи это 0,69 г (Таблица 4).
- Пшеница (корневая часть). Представлены рубрики с выжившей вегетативной (надземной) частью к концу эксперимента, т.е. на 25-й день полива.
Растения в грунте N/N и N/R, а также стресс-почве N/N и стресс-почве N/R погибли соответственно на 1-й, 1-й, 1-й и 1-й неделе испытания.
Таблица 5.
Убыль корневой массы пшеницы в ходе эксперимента.
|
Дни экспериментального полива |
Грунт O /R |
Стресс-грунт O/R |
|
0 день |
0,15 |
0,15 |
|
5 день |
0,12 |
0,09 |
|
10 день |
0,08 |
0,05 |
|
15 день |
0,06 |
0,03 |
|
20 день |
0,04 |
0,02 |
|
25 день |
Ʃ 0.03
O /R 0,02/0,01 |
Ʃ 0,01
O /R ? |
График зависимости убыли корневой массы пшеницы (г)—Оу от времени (дни)—Ох.
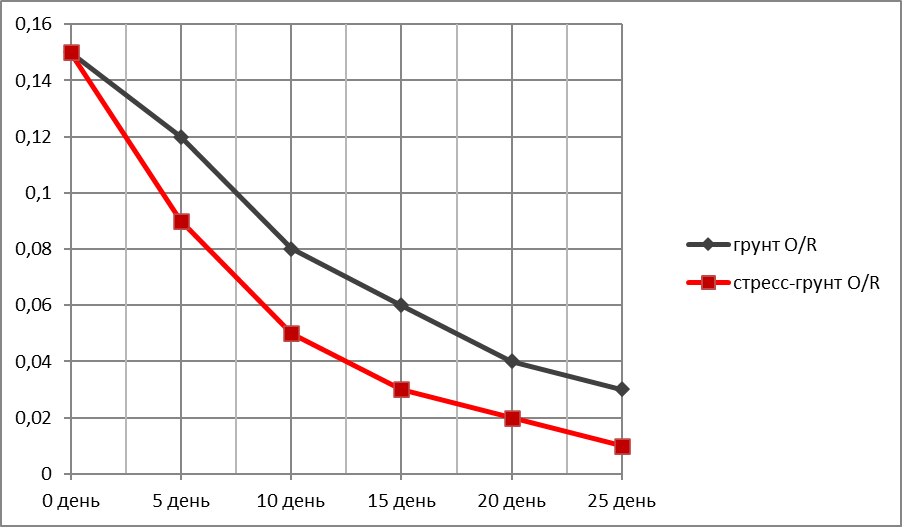
За исходную массу корневой системы (день 1) бралась масса корней трёх здоровых пророщенных/укоренившихся растений после 1-й фазы испытаний. Для пшеницы это 0,15 г (Таблица 5).
Таблица 6.
Убыль массы вегетативной части пшеницы в ходе эксперимента.
Представлены образцы, сохранившие живой вегетативную часть все 25 дней опыта.
Растения в грунте N/N и N/R и стресс-почве N/N и стресс-почве N/R погибли соответственно на 1-й и 2-й ; 1-й и 1-й неделе испытания.
|
Дни экспериментального полива |
Грунт O /R |
Стресс-грунт O/R |
|
0 день |
0,57 |
0,57 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5 день |
0,43 |
0,38 |
|
10 день |
0,31 |
0,26 |
|
15 день |
0,22 |
0,16 |
|
20 день |
0,14 |
0,09 |
|
25 день |
0,08
|
0,05
|
График зависимости убыли массы надземной (вегетативной) части пшеницы (г)—Оу от времени (дни)—Ох.
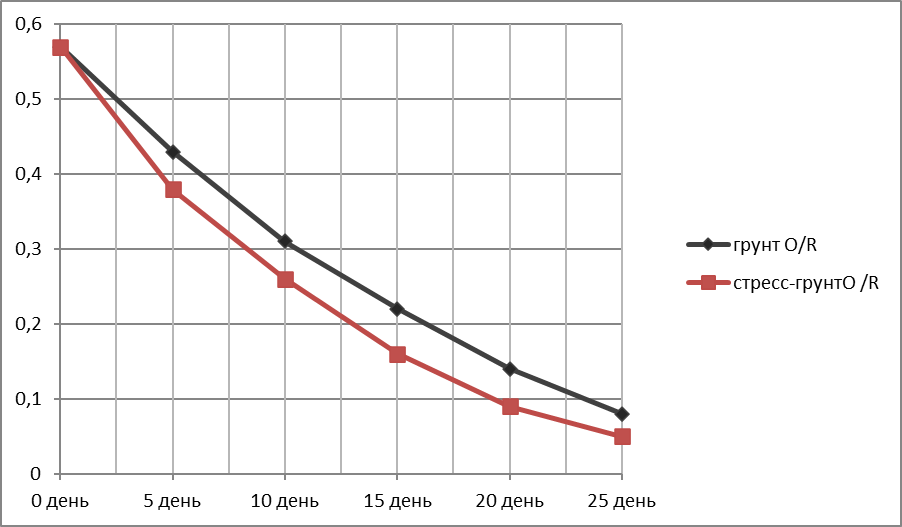
За исходную массу вегетативной части (день 1) бралась масса трёх здоровых средних по размеру пророщенных растений в чернозёме без стресса разделения корней после 1-й фазы испытаний. Для пшеницы это 0,57 г (Таблица 6).
Результаты практической части эксперимента:
- Растения, выдержавшие все фазы испытания: кукуруза, пшеница, рожь—являются засухоустойчивыми культурами по энциклопедическим сведениям.
- Ни одно растение не стало адекватно развиваться в условиях стресса, но каждый вид растения боролся со стрессом во времени по-разному.
- При анализе борьбы растений со стрессом в чернозёме выявлено, что растения, имея только стресс в виде разделения корней по разным ёмкостям, лучше выживают при факторе недостаточного полива, в группе N/R и O/R, вопреки представлениям о том, что лучший результат ожидаем в группе N/N с достаточным типом полива.
Т.е., в ёмкостях, где не включается фактор борьбы за воду— растения гибнут значительно быстрее (на первой же неделе эксперимента).
По-видимому, это связано с тем, что борьба за воду, для растений является филогенетически важнейшим и древнейшим механизмом. Активизируясь, путём трансформации сотен-тысяч защитных генов, по этой же цепочке запускаются процессы борьбы растения и с другими факторами стресса.
Если же механизм борьбы растения за воду не включается, со стрессом другой модальности (разделение корней) растения справляются значительно хуже.
- Растения, выдержавшие испытания в виде разделения корней в условиях именно недостаточного полива, успешнее выдерживают и полифакторный стресс (разделение корней + обеднённая засоленная эрозированная почва с примесью тяжёлых металлов), что подтверждает тезис о том, что именно при включении механизма борьбы за воду, одновременно запускаются процессы борьбы со стрессом другой модальности, в том числе полифакторным.
- При взвешивании корневой системы в каждой паре ёмкостей финальной части эксперимента, выявлено, что корневая система угасала быстрее в ёмкостях с более неадекватным типом полива. В паре N/R растение большей частью брало воду из ёмкости с нормальным типом полива, а в паре O/R—предпочитало запитываться из ёмкости О с экстремально низким, однако ежедневным типом полива, чем предпочитать риск из ёмкости R c адекватным по объёму, но редким во времени типом полива (см. таблицу убыли корневой массы во времени). Т.е. растения, участвовавшие в эксперименте и дошедшие до его финала: кукуруза, рожь и пшеница не склонны рисковать и выбирают более надёжный во времени источник полива, пусть даже он неадекватен по объёму.
- Нами замечено, что графики убыли массы корней и вегетативной части растений в условиях стресса— схожи с финальной частью зоны угнетения в биологической схеме действия экологического фактора на жизнедеятельность организмов (рис. 10), что логично, однако, эта зависимость не в полной мере характеризует интенсивность борьбы растения со стрессом, не имея количественного (математического) отражения.
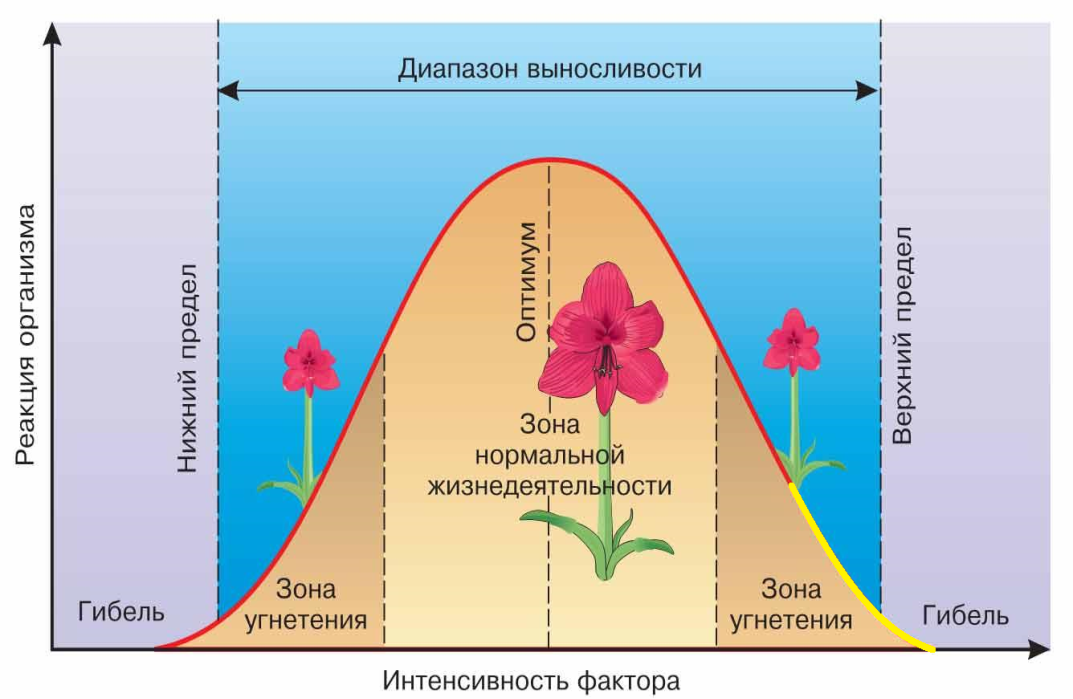
Рис.10. Биологическая схема действия экологического фактора на жизнедеятельность организмов.
- В поисках адекватного эквивалента для математического выражения зависимости борьбы растения со стрессом, нами замечено, что убыль массы (как корневой, так и вегетативной) с течением времени так же схожа с графиком закона радиактивного распада (рис.11), а значит, коэффициент борьбы растений со стрессом, можно высчитать и представить полноценной математической величиной, приняв за него эквивалент времени полураспада ядер радиоактивного элемента.
- К слову, данную зависимость не раз замечали в учёном мире, применимо к другим областям жизни, например, в физической химии для описания реакций убыли вещества первого порядка, а так же для описания законов демографии при убыли народонаселения [3,с.30-31].
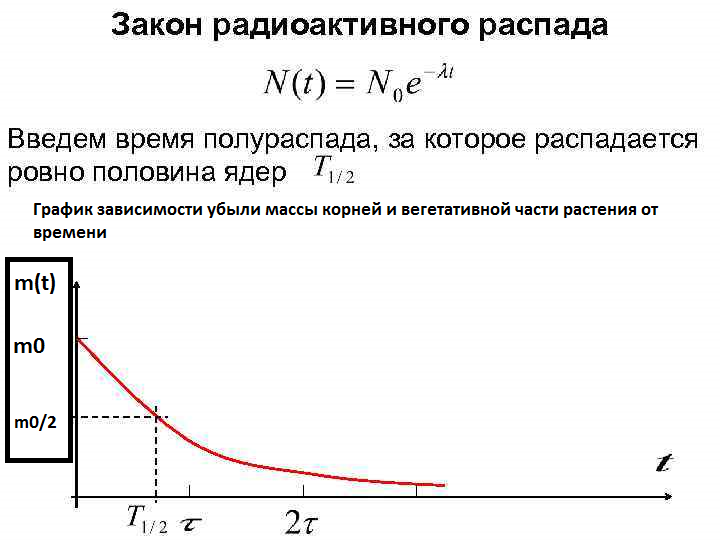
Рис. 11. Закон радиоактивного распада (вместо убыли массы ядер—убыль массы растения во времени)
Перерисовав графики на миллиметровую бумагу (рис.12), высчитан эквивалент времени полураспада для измерения стрессоустойчивости растений, конечные результаты которого отражены в наглядном виде для корневой и вегетативной части соответственно (Таблица 7, Таблица 8).
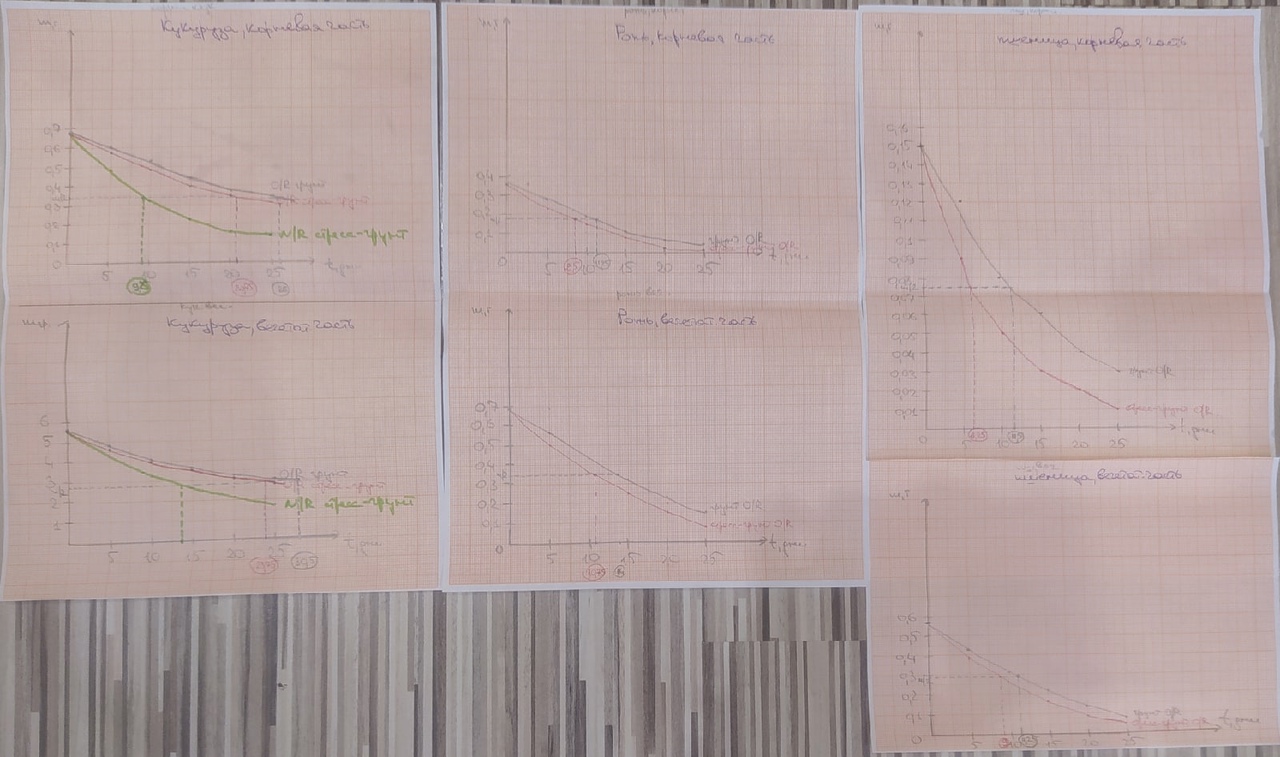
Рис. 12. Графики закона радиоактивного распада для расчёта убыли массы растений во времени на миллиметровой бумаге.
Таблица 7.
Эквивалент времени полураспада для измерения стрессоустойчивости растений для корневой части.
|
Т1/2, дни
|
Грунт, вид полива Опрыскивание/Риск |
Стресс-грунт, вид полива Опрыскивание/Риск |
Стресс-грунт, вид полива Норма/Риск |
|
Кукуруза |
26 |
20,75 |
9,25 |
|
Рожь |
11,25 |
8,5 |
-- |
|
Пшеница |
11,5 |
6,25 |
- |
Таблица 8.
Эквивалент времени полураспада для измерения стрессоустойчивости растений для вегетативной части.
|
Т1/2, дни
|
Грунт, вид полива Опрыскивание/Риск |
Стресс-грунт, вид полива Опрыскивание /Риск |
Стресс-грунт, вид полива Норма/Риск |
|
Кукуруза |
30,5 |
23,75 |
13,75 |
|
Рожь |
14 |
10,75 |
-- |
|
Пшеница |
11,25 |
9 |
-- |
Таким образом, мы приходим к выводу о том, что для наглядного отражения стрессоустойчивости растений можно опираться на физический закон радиоактивного распада.
Чем больше цифра полученного коэффициента для определенного вида растения, тем более стрессоустойчивым оно является. Изотопная хронология, или радиоуглеродное датирование,--это способ подсчёта времени, прошедшего с тех пор, как живая материя стала неживой. В течение жизни растения количество углерода-14 (6С14) поддерживается постоянным за счёт постоянного пополнения из атмосферного воздуха. В атмосфере углерод-14 появляется за счёт воздействия космических лучей. После смерти живого организма поступление углерода-14 прекращается, и его излучение постепенно ослабевает. Возраст сохранившегося органического материала можно определить радиоуглеродным методом. Спад активности, т.е. число распадов в секунду, для некоторого радиоактивного вещества, в том числе и для углерода-14, это и есть физический график радиоактивного распада! [4,с.3-4].
В заключение хотелось бы сказать, что законы мира универсальной природы, что в последнее время всё больше подтверждает квантовая физика, и привести цитату В.В. Соколова: « К физике принадлежит учение о небесах, о стихиях (элементах) и их движении, о возникновении и уничтожении, о влиянии небес на погоду (метеорология), о минералах, растениях, животных, о душе и её способностях. Медицина, астрология, учение о талисманах, алхимия, толкование снов и волшебства представляют собой лишь разновидности прикладной физики».
Не случайны и в гимне МФТИ такие слова: «Только в физике соль…»!
- Борисова. Г. Г., Малева М.Г., Чукина Н.В. Лекции по фитофизиологии Уральского государственного университета им. Горького. Растение и стресс [Электронный ресурс]. Екатеринбург. 2008. С.6-8. URL: http://www.elar.urfu.ru/bitstream/10995/1580.
- Доанг Хоанг Жанг, Тохтарь В.К. Исследование засухоустойчивости перспективных для интродукции видов растений// БелГУ Научные ведомости. Естественные науки, 2011. №9 (104). С.43-46.
- Стасенко А. Радиоактивный распад, банковский процент и др.// Квант. 2018. N8. С.30-31.
Научные высказывания #99